Article
깃털 성감별 상업용 토종 실용닭 생산
손시환1,†
,
조은정2,
이슬기3,
이준호4,
장수용4,
용귀중5
Production of Feather-Sexing Korean Native Commercial Chickens
Sea Hwan Sohn1,†,
Eun Jung Cho2,
Seul Gy Lee3,
Junho Lee4,
Suyong Jang4,
Kwijoong Yong5
Author Information & Copyright ▼
1Professor, Division of Animal Bioscience and Integrated Biotechnology, Gyeongsang National University, Jinju 52725, Republic of Korea
2Researcher, Division of Animal Bioscience and Integrated Biotechnology, Gyeongsang National University, Jinju 52725, Republic of Korea
3Graduate Student, Division of Animal Bioscience and Integrated Biotechnology, Gyeongsang National University, Jinju 52725, Republic of Korea
4Student, Division of Animal Bioscience and Integrated Biotechnology, Gyeongsang National University, Jinju 52725, Republic of Korea
5Manager, Hanhyup Pure Line Co. Ltd., Danyang 27001, Republic of Korea
© Copyright 2024, Korean Society of Poultry Science. This is an Open-Access article distributed under the terms of the
Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0/) which permits
unrestricted non-commercial use, distribution, and reproduction in any
medium, provided the original work is properly cited.
Received: May 06, 2024; Revised: May 20, 2024; Accepted: May 21, 2024
Published Online: Jun 30, 2024
적 요
병아리의 성 감별 방법으로 깃털 감별법이 상업적으로 가장 널리 이용되고 있다. 그러나 깃털 감별법이 산업적으로 실용화되기 위해서는 종계의 기초 계군 내 조우성과 만우성유전자가 존재하고, 깃털 감별이 될 수 있는 종계 계통을 조성하여야 하고, 깃털 감별로서 병아리의 성 판별율이 정확하여야 한다. 따라서 본 연구에서는 상업용 토종닭의 깃털 감별을 위한 자가 성감별 종계 계통 조성 방법과 조성된 종계로써 생산한 상업용 병아리들의 깃털 성 판별의 유효성을 소개하고자 하였다. 깃털 감별용 종계 조성을 위한 기초 계군은 조우성 개체와 만우성 개체가 혼재되어 있었으며, 만우성유전자의 빈도가 0~0.205 정도로 추정되었다. 병아리의 깃털 자가 성감별을 위한 계통 조성으로 원종계의 부 계통은 암수 공히 조우성(kk)으로 고정하고, 모 계통의 부는 만우성 동형접합체(ZKZK)를 가진 계군으로, 모는 조우성(ZkW)을 가진 계군으로 조성하였다. 따라서 부계 종계는 조우성(ZkZk), 모계 종계는 만우성(ZKW)으로 되어 이들 간 교잡으로 생산된 병아리들은 수컷은 만우성(ZKZk), 암컷은 조우성(ZkW)을 가지므로 병아리의 깃털 형태로써 암수를 감별할 수 있었다. 또한 생산된 실용 병아리에 대한 깃털 감별의 유효성을 살펴보고자 생산 병아리 중 1,000수를 표본으로 항문 감별과 깃털 감별 간의 감별 일치도를 조사한 결과 일치율은 93.1%로 나타났다.
ABSTRACT
The feather-sexing method is widely used commercially for chick sex identification. However, for feather-sexing to be industrially practical, the early-feathering (EF) and late-feathering (LF) genes must existed within the foundation stock, a suitable feather-sexing lines must be established, and the accuracy of sex identification by feather-sexing must be ensured. Therefore, this study introduces the method of constructing the Korean native chickens (KNC) feather-sexing lines using EF and LF genes and evaluates the effectiveness of feather sex determination on commercial chicks produced from the constructed KNC lines. The results showed that both EF and LF chickens existed within the foundation stock, with the frequency of LF genes ranging from 0 to 0.205. In feather-sexing line establishment, the paternal strain of the grandparent stock (GPS) was fixed as EF (kk) for both sexes, while the maternal strain was composed of males with LF homozygotes (ZKZK) and females with EF (ZkW). Thus, in the parent stock (PS), male breeder had EF (ZkZk) and female breeder had LF (ZKW), resulting in chicks produced from their crosses having LF (ZKZk) for males and EF (ZkW) for females, allowing sex determination based on feather development. Additionally, to evaluate the effectiveness of feather-sexing for the produced commercial chicks, a study was conducted on 1,000 samples of the produced chicks to investigate the concordance between vent-sexing and feather-sexing, showing a matching rate of 93.1%.
Keywords: 한국토종실용닭; 깃털감별계통 조성; 조우성; 만우성
Keywords: Korean native commercial chickens; feather-sexing line construction; early-feathering; late-feathering
서 론
병아리의 성 감별은 가금 산업의 경제적 측면에서 매우 중요하다. 병아리의 해부 생리적 특성상 발생 직후 외관상으로 암수의 구분이 불가능하기 때문에 병아리의 암수를 식별하고자 하는 다양한 연구들이 수행되고 있다. 최근에는 부화 전 수정란 상태에서 배자의 성을 판별하고자 하는 연구들이 시도되고 있지만 아직은 이러한 방법들이 산업적으로 적용되기에는 여러가지 어려움이 있다(Weissmann et al., 2013; Webster et al., 2015; Clinton et al., 2016; Galli et al., 2018; Jia et al., 2023). 따라서 부화 직후 병아리의 성을 감별하는 기술들이 보편적으로 활용되고 있는데, 이러한 방법 중 지금까지 가장 대표적인 성 감별 방법은 발생 병아리의 생식 돌기의 형태적 차이로써 성을 판별하는 항문돌기감별법(Martin, 1934)과 반성 유전을 이용한 깃털 감별법(Somes, 1969; Warren, 1976; Gawron and Smyth, 1980; Bacon et al., 1988; Campo, 1991; Bitgood, 1999; Sohn et al., 2012)이 있다. 가장 보편적으로 이용되는 항문돌기감별법은 정확도가 높은 반면 전문 감별사의 감별 기술에 의존함으로써 감별 기술을 숙련하는 데 오랜 시간이 필요하고, 시간적, 경제적 부담이 큰 단점이 있다. 따라서 최근에는 반성 유전을 이용한 성 판별이 상업적으로 실용화되어 있는데, 특히 닭의 깃털 발생 속도와 연관된 조우성(early-feathering)과 만우성(late-feathering)을 이용한 깃털 감별법을 가장 널리 이용하고 있다. 이는 모계를 만우성(ZKW)으로 하고, 부계를 조우성(ZkZk) 닭으로 하였을 때 암평아리는 조우성(ZkW), 수평아리는 만우성(ZKZk)이 되어 깃털 발육 형태로써 성을 판별할 수 있는 방법이다(Somes, 1969; Iraqi and Smith, 1995; Bang et al., 2018; Kwon et al., 2021). 그러나 깃털 감별법이 산업적으로 실용화되기 위해서는 종계의 기초 계군 내 조우성과 만우성 유전자가 존재하여야 하고(Sohn et al., 2012), 조우성과 만우성 개체의 정확한 식별 방법이 확립되어야 하며(Siegel et al., 1957; Aviagen Brand, 2018; Bang et al., 2018), 깃털의 조만성에 따른 개체 간 생산능력의 차이도 없어야 하고(Somes, 1970; Lowe and Garwood, 1981; Dunnington and Siegel, 1986; Lowe and Merkley, 1986; O’Sullivan et al., 1991; Fotsa et al., 2001; Nahashon et al., 2004; Khosravinia, 2008; Sohn et al., 2013, 2021; Kim et al., 2019), 무엇보다 깃털 감별이 될 수 있는 종계 계통이 조성되어야 한다(Kwon et al., 2021). 더불어 깃털 감별로서 성 판별율의 정확성이 높아야 한다. 따라서 이러한 조건들이 완비되었을 때 병아리의 상업적 성 감별이 가능한데 현재 실용계 생산을 위한 수입 종계 대부분은 깃털 감별이 가능한 계통으로 조성된 상태이나 토종닭의 경우는 최근에 자가 깃털 감별 토종 종계를 조성하고 시험적으로 상업용 병아리의 암수 감별을 시도하고 있는 실정이다. 따라서 본 연구에서는 한국토종닭의 깃털 감별을 위한 자가 성감별 종계 계통 조성 방법과 이들 종계로부터 생산된 상업용 시판 병아리들의 깃털 성 판별의 유효성을 제시하고자 하였다.
재료 및 방법
1. 공시동물 및 사양관리
본 연구에 이용한 토종닭의 기초 계군은 ㈜한협원종이 보유한 순계(pure line; PL)로써 이를 이용하여 깃털 감별이 가능한 4개 계통의 원종계(grand parent stock; GPS) 및 종계(parent stock; PS)를 조성하였다. 이들이 보유한 순계는 1962년부터 도입한 Cornish, White Plymouth Rock, Barred Plymouth Rock, Patridge Rock, Rhode Island Red 및 Hampshire 종을 기초로 하여 10개의 토착 계통으로 고정하여 조성된 계군이다. 순계 및 원종계의 사양 관리는 강제 환기 및 자동 온도조절시스템이 완비된 종계사에서 단사 케이지(580 cm2/1수)로 개체 사육하였다. 사료 급여는 사육 단계별로 종계 사양관리 지침에 따라 시판용 초이사료, 병아리 육성기 사료, 산란기 사료를 급여하고 3주령 이후부터 체중별 상위, 중위, 하위 그룹으로 나누어 ㈜한협원종의 육용 종계 권장 급여량으로 제한 급여하였다. 점등 관리는 종계 표준 점등프로그램에 따라 실시하였고, 사양 기별 백신 접종 및 위생관리는 ㈜한협원종의 표준 백신 접종프로그램에 따라 수행하였다.
2. 조우성과 만우성 깃털 식별방법
1) 깃털 형태에 따른 조우성과 만우성의 식별
깃털 형태에 의한 조우성과 만우성의 식별은 발생 직후 날개 깃의 형태 및 5일령 이후 꼬리 깃의 발육 양상으로 감별하였다. 식별 방법은 Bang et al.(2018)이 제시한 병아리의 깃털 발생 양상을 기준으로 발생 직후 날개 깃의 주익우(primaries)와 부익우(coverts)의 형태 및 길이의 차이로 구분하였는데, 주익우가 상대적으로 길고 부익우와 길이 차이가 현저하면 조우성으로 분류하고, 주익우가 상대적으로 짧고 주익우와 부익우의 길이 차이가 거의 없으면 만우성으로 분류하였다. 한편 5일령 이후 꼬리 깃의 발육 양상으로 조우성과 만우성의 식별 방법은 조우성의 경우 꼬리 깃의 돌출이 명확하게 나타나고, 만우성은 이러한 양상이 나타나지 않은 것으로 분류하였다(Fig. 1).
Fig. 1.
Feather phenotypes of early- and late-feathering chicks. On day one, early-feathering chicks exhibit long primaries and coverts, with a distinct difference in length between primaries and coverts. Late-feathering chicks have shorter primaries and coverts with similar lengths. At 5 days old, the tail feathers of early-feathering chicks are long and distinct, whereas those of late-feathering chicks are short and unclear.
Download Original Figure
2) PCR을 이용한 조우성과 만우성의 식별
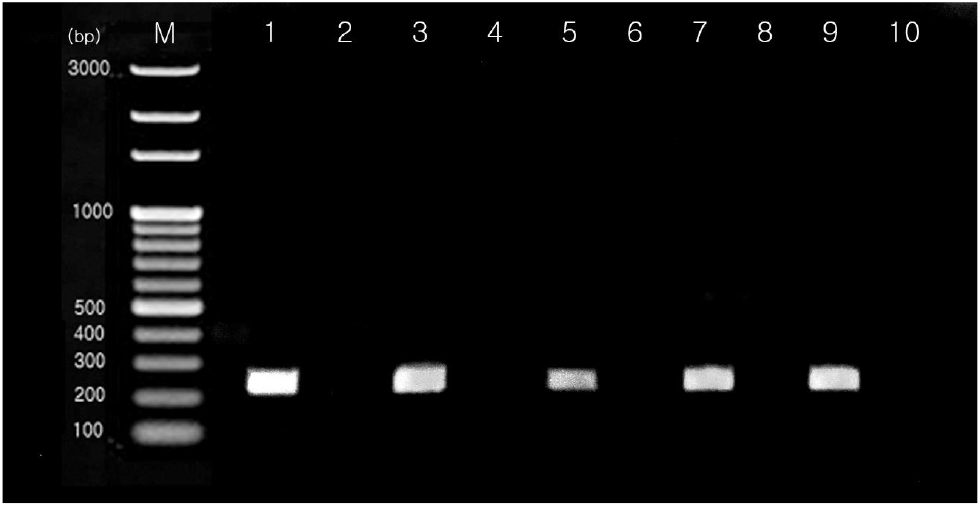
조우성과 만우성의 개체 식별을 위하여 만우성 특이 DNA primer를 이용하여 polymerase chain reaction(PCR)을 실시하였다. 만우성 특이 DNA primer의 제작 및 PCR의 방법은 Bang et al.(2018)이 제시한 방법과 동일하게 수행하였다. 공시한 개체들의 혈액으로부터 PCR의 반응물은 1.5% agarose gel로 전기 영동한 후 만우성유전자의 band 유무로 조우성과 만우성 개체를 판단하였다(Fig. 2). 본 시험에 관련된 닭의 관리 및 취급은 경상국립대학교 동물실험윤리위원회(IACUC, No. 2020-5)의 규정을 준수하여 시행하였다.
Fig. 2.
PCR results using the chicken K specific gene primers. M represents the 100 bp size marker. Late-feathering (LF) chickens in lanes 1, 3, 5, 7, and 9 demonstrate the presence of 230 bp K specific gene products of the same size. No products are observed in early-feathering (EF) chickens in lanes 2, 4, 6, 8, and 10.
Download Original Figure
3. 종계 계통 조성 및 유전자형 식별을 위한 검정 교배
깃털 감별 계통 조성을 위하여 10개의 토착 순계 계통에 대해 계통 내 조우성과 만우성 개체의 분포 빈도를 조사하고 생산 능력 및 조만성 분포 양상을 기초로 하여 적합한 계군을 선정하였다. ㈜한협원종이 보유한 10개의 토착 순계 계군 중 4개의 계군을 원종계 계통(GPS)으로 선정하고 이들 간의 교잡으로 종계 계통(PS)을 설정하였다. 깃털 자가 성감별 병아리 생산을 위한 계통 조성 체계는 Fig. 3과 같다. GPS 부 계통의 부계(父系)인 A계통은 유전자형이 ZkZk로 고정된 수컷 170수를, 모계(母系)인 B계통은 ZkW인 암컷 692수를 선발하였고, 모 계통의 부계인 C계통은 ZKZK인 수컷 207수, 모계인 D계통은 ZkW인 암컷 2,054수를 선발 공시하였다. GPS 모 계통의 부(父)는 만우성 동형접합체(ZKZK)를 가져야함으로 이의 선발을 위해 순계(PL) 만우성 수컷 176수를 대상으로 수컷당 암컷 6수와 검정 교배를 수행하였고, 이들로부터 생산된 자손들의 조우성과 만우성의 발생 양상을 분석하여 만우성 동형접합체를 가진 부계를 선발하였다.
Fig. 3.
Breeding system of line construction for the production of feather-sexing chicks. GPS; Grandparent stock, PS; Parent stock, CC; Commercial chicks, EF; Early-feathering, LF; Late-feathering.
Download Original Figure
4. 실용계의 깃털 자가성감별 유효성 검정
병아리의 깃털 감별을 위해 모계는 만우성(ZKW)으로 하고, 부계는 조우성(ZkZk)으로 하였을 때 생산되는 수평아리는 만우성, 암평아리는 조우성이 된다. 따라서 이와 같은 이론적 감별의 유효성을 확인하기 위하여 공시된 종계의 교잡으로 생산된 병아리 중 표본으로 5개 농장 1,000수에 대해 항문 감별과 깃털 감별 간 감별의 일치도를 검정하였다.
결과 및 고찰
1. 기초 계군의 조만성 분포 양상과 유전자 빈도
깃털 감별 계통 조성을 위한 4개의 기초 계군(PL) 내 조우성과 만우성 개체의 분포 빈도를 조사하고, 이를 바탕으로 만우성 유전자(K)의 유전자 및 유전자형 빈도를 추정하여 이의 결과를 Table 1에 제시하였다. 분석 결과 A, B, C 세 계군은 조우성 개체와 만우성 개체가 혼재되어 있으나 D계군은 조우성 개체만 존재하는 것으로 나타났다. 본 집단이 Hardy-Weinberg 평형상태에 있다는 가정하에 조만우성 개체의 분포 빈도로써 유전자 및 유전자형 빈도를 추정한 결과 만우성유전자(K)의 빈도가 D계군은 0인 반면 나머지 세 계군은 0.062~0.205 정도로 나타났다. 집단 내 만우성 유전자의 분포 빈도는 품종에 따라 큰 차이를 보이는데 국내 유전자원으로 보존 중인 토종닭 10품종에서 토종 흑색코니시종, 토종 로드종 및 한국재래닭 적갈색종에만 조우성과 만우성의 유전자가 혼재되어 있고, 토종 갈색코니시종, 백색레그혼종, 오골종, 재래황갈종, 재래회색종, 재래백색종 및 재래흑색종은 집단 내 만우성 유전자가 없는 것으로 나타났다(Sohn et al., 2012). 한편, 상업용 실용토종닭에서 만우성 개체가 약 13%정도 나타난다는 보고(Bang et al., 2018)에 따라 이들의 종계 집단 내 만우성유전자가 존재하는 것으로 판단되어 깃털 감별 계통 조성의 계기가 되었다. 따라서 깃털감별계통 조성을 위해서는 반드시 기초 집단 내 조우성과 만우성 개체가 혼재되어 있어야 하고 만우성 유전자의 분포 빈도가 일정 수준은 되어야 한다. 그러므로 본 기초 집단의 경우 깃털감별계통 조성을 위한 계군별 만우성 유전자가 존재하고, 이의 빈도 또한 계통 조성에 충분한 빈도로 나타나 깃털 자가 성감별을 위한 종계 계통 조성이 가능한 것으로 판단되었다.
Table 1.
Distribution of late- and early-feathering chickens, and gene and genotype frequencies for K gene in feather-sexing foundation stocks
| Stocks |
Tested chickens |
No. chickens |
Gene frequency |
Genotype frequency |
| Late-feathering |
Early-feathering |
K
|
k
|
KK
|
Kk
|
kk
|
| A |
830 |
100 |
730 |
0.062 |
0.938 |
0.004 |
0.116 |
0.880 |
| B |
1,220 |
321 |
899 |
0.142 |
0.858 |
0.020 |
0.244 |
0.736 |
| C |
1,128 |
415 |
713 |
0.205 |
0.795 |
0.042 |
0.326 |
0.632 |
| D |
2,470 |
0 |
2,470 |
0.000 |
1.000 |
0.000 |
0.000 |
1.000 |
Download Excel Table
2. 깃털 자가 성감별 종계 계통 조성
만우성유전자를 이용하여 병아리의 자가 성감별을 하기 위해서 원종계(GPS)의 부 계통은 암수 공히 조우성(kk)으로 고정되어야 하고, 모 계통의 부(父)는 만우성 동형접합체(ZKZK)를 가진 계군으로 고정되어야 하며, 모 계통의 모(母)는 조우성이나 만우성 어떠한 개체라도 상관이 없다. 이러한 원종계 교배조합에 따라 종계(PS)의 부(父)는 조우성(ZkZk), 모(母)는 만우성(ZKW)을 가지게 됨으로 종계 간 교잡으로 생산되는 실용계(CC)의 암평아리는 조우성, 수평아리는 만우성이 되어 자가 성감별이 가능하게 된다(Fig. 3). 따라서 본 종계의 계통 조성에서 원종계 부 계통의 부계(父系)인 A계통은 ZkZk로 고정한 수컷으로, 모계(母系)인 B계통은 ZkW로 고정한 암컷으로 조성하고, 모 계통의 부계인 C계통은 ZKZK인 수컷으로, 모계인 D계통은 ZkW의 유전자 형을 지닌 암컷으로 조성하였다. 원종계 모 계통의 부(父)인 C계통은 만우성 동형접합체(ZKZK)를 가져야함으로 이의 선발을 위해 해당 계통의 만우성 순계(PL) 수컷 176수를 대상으로 수컷 1수당 조우성 암컷 6수를 검정 교배하고 생산된 모든 자손들의 조우성과 만우성을 조사하였다. 검정 결과, 조우성 자손을 한 마리도 생산하지 않은 검정 개체는 10수로 이들을 만우성 동형접합체(ZKZK)를 가진 개체로 추정하였고, 나머지 166수는 만우성 이형접합체(ZKZk)를 가진 개체로 판단하였다. 이는 C축군의 유전자 및 유전자 빈도 추정시 KK의 빈도가 4.2%로 추정된 것과 거의 비슷한 분포 양상인 것으로 나타났다. 따라서 C계통의 원종계 조성은 만우성 동형접합체를 가진 것으로 판단되는 수컷 순계 10수를 이용하여 만우성 암컷 개체와 교잡하여 ZKZK인 수컷 207수를 생산하였다.
3. 실용계의 깃털 자가 성감별 유효성 검정
원종계 A와 B계통의 교잡으로 부계 종계 AB(ZkZk)를 조성하고, C와 D계통의 교잡으로 모계 종계 CD(ZKW)를 조성하여 이들 간 교잡으로 실용계를 생산하였다. 따라서 생산된 실용계 병아리(ABCD)들은 수컷은 만우성(ZKZk), 암컷은 조우성(ZkW)을 가지게 됨으로 병아리의 깃털 형태로써 암수를 쉽게 구별할 수 있다. 이러한 종계 조성 체계에 따라 한 해 동안 총 176,100수의 종계(PS)를 생산하였다. 이들 종계로써 생산된 실용 병아리에 대한 깃털 감별의 유효성을 살펴보고자 이 중 1,000수를 표본으로 5개 농장에서 각기 항문 감별과 깃털 감별 간의 감별 일치도를 조사하였다. 분석 결과, Table 2와 같이 농장 별로 91.5~94.0%정도의 감별 일치율을 보이고, 전체 1,000수 중 항문 감별과 깃털 감별 간 93.1%가 일치하였다. 이는 실험적으로 수행한 깃털 자가 성감별의 결과보다는 다소 낮은 성적으로 Kwon et al.(2021)이 한국재래닭 적갈색종으로 조성한 조우성 부계와 만우성 모계로부터 생산한 총 140수의 병아리 중 깃털 감별로서 두 마리만 감별 오차가 나타나 98.6%의 감별율을 보고하였는데 두 마리 모두 PCR 검증 결과 만우성 개체를 조우성 개체로 잘못 식별한 경우이다. 본 연구에서 감별 일치율은 항문 감별과 깃털 감별 간의 비교로 실제 개체의 성과 감별의 일치율을 나타내는 것이 아니다. 이는 항문 감별에서도 감별의 오차가 발생할 수 있고, 깃털 감별에서도 감별의 오차가 발생할 수 있음으로 실제 개체의 성과의 감별에 있어 어떠한 방법이 더욱 정확한지는 추후 실험이 필요한 과제이다. 그럼에도 불구하고 본 비교에서 나타난 차이는 대부분 조우성과 만우성 개체의 깃털 식별 오차로 판단된다. 만약 깃털 자가 성감별을 위한 종계의 계통 조성에 오류가 있었다면 상당수의 실용계가 성 판별 오차로 나타나기 때문이다. 개체에 따라 외관상 조우성과 만우성 깃털 형태의 차이를 식별하는 데 구분이 모호한 경우가 다소 있어 대개 만우성 개체를 조우성 개체로 구분하는 경우가 많다. 조우성과 만우성의 식별이 바뀜으로 성 판별이 달라지기 때문에 조만성 깃털 식별의 정확도가 매우 중요하다.
Table 2.
The concordance rate of sex identification results between vent-sexing and feather-sexing method
| Farms |
Vent-sexing |
Feather-sexing |
Matching rate b/w vent- and feather-sexing |
| Male |
Female |
Total |
Male |
Female |
Total |
| G farm |
99 |
101 |
200 |
82 |
118 |
200 |
91.5% |
| C farm |
98 |
102 |
200 |
85 |
115 |
200 |
93.5% |
| K farm |
98 |
102 |
200 |
86 |
114 |
200 |
94.0% |
| HH farm |
98 |
102 |
200 |
83 |
117 |
200 |
92.5% |
| HR farm |
98 |
102 |
200 |
86 |
114 |
200 |
94.0% |
| Total |
491 |
509 |
1,000 |
422 |
578 |
1,000 |
93.1% |
Download Excel Table
사 사
본 논문은 농촌진흥청 2024년도 농업생명자원 관리기관 운영과제(RS-2024-00424820)의 지원으로 수행되었음.
REFERENCES
Bacon LD, Smith E, Crittenden LB, Havenstein GB 1988 Association of the slow feathering (
K) and an endogenous viral (
ev21) gene on the Z chromosome of chickens. Poult Sci 67(2):191-197.



Bang MH, Cho EJ, Cho CY, Sohn SH 2018 Study on the characteristics of feather developing pattern and morphology in early- and late-feathering Korean native chickens. Kor J Poult Sci 45(3):155-165.
Clinton M, Nandi S, Zhao D, Olson S, Peterson P, Burdon T, McBride D 2016 Real-time sexing of chicken embryos and compatibility with
in ovo protocols. Sex Dev 10(4):210-216.
Dunnington EA, Siegel PB 1986 Sex-linked feathering alleles (
K,
k+) in chickens of diverse genetic backgrounds. 1. Body temperatures and body weights. Poult Sci 65(2):209-214.
Fotsa JC, Mérat P, Bordas A 2001 Effect of the slow (
K) or rapid (
k+) feathering gene on body and feather growth and fatness according to ambient temperature in a Leghorn x brown egg type cross. Genet Sel Evol 33(6):659-670.
Jia N, Li B, Zhao Y, Fan, S, Zhu J, Wang H, Zhao W 2023 Exploratory study of sex identification for chicken embryos based on blood vessel images and deep learning. Agriculture 13(8):1480.
Khosravinia H 2008 Effect of the slow (
K) or rapid (
k+) feathering gene on growth performance and skeletal dimensions of broiler chickens selected for cut up carcass value. Res J Poult Sci 2(1):9-14.
Kim KG, Cho EJ, Choi ES, Kwon JH, Jung HC, Sohn SH 2019 Comparison of production performances between early- and late-feathering chickens in parent stocks of Korean native chicken. Kor J Poult Sci 46(4):279-286.
Kwon JH, Choi ES, Sohn SH 2021 Establishment of Korean native chicken auto-sexing lines using sex-linked feathering gene. Kor J Poult Sci 48(1):41-50.
Lowe PC, Garwood VA 1981 Independent effects of K and k+ alleles and maternal origin on mortality and performance of crossbred chickens. Poult Sci 60(6):1123-1126.
Lowe PC, Merkley JW 1986 Association of genotypes for rate of feathering in broilers with production and carcass composition traits: effect of genotypes, sex, and diet on growth and feed conversion. Poult Sci 65(10):1853-1858.
Martin JH 1934 Review the “Sexing baby chicks by Masui and Hashimoto”. Poult Sci 13(3):190.
Nahashon SN, Bartlett J, Smith EJ 2004 Effect of the late-feathering or early-feathering genotypes on performance and carcass traits of broiler chickens. Live Prod Sci 91(1):83-94.
O’Sullivan NP, Dunnington EA, Siegel PB 1991 Growth and carcass characteristics of early- and late-feathering broilers reared under different feeding. Poult Sci 70(6):1323-1332.
Siegel PB, Mueller CD, Craig JV 1957 Some phenotypic differences among homozygous, heterozygous, and hemizygous late feathering chicks. Poult Sci 36(2):232-239.
Sohn SH, Choi ES, Cho EJ, Kim BG 2021 Effects of early- and late-feathering phenotypes on growth performance and mortality in Korean native commercial chickens. Kor J Poult Sci 48(4):177-184.
Sohn SH, Kim NY, Park DB, Song HR, Cho EJ, Choi SB, Heo KN, Choi HC 2013 Influence of early- and late-feathering phenotype on productive performance in the feather-sexing strains of Korean Native Chicken. Kor J Poult Sci 40(3):263-270.
Sohn SH, Park DB, Song HR, Cho EJ, Kang BS, Suh OS 2012 Genotype frequencies of the sex-linked feathering and their phenotypes in domestic chicken breeds for the establishment of auto-sexing strains. J Anim Sci Tech 54(4):1-10.
Somes RG 1969 Delayed feathering, a third allele at the
K locus of the domestic fowl. J Hered 60(5):281-286.
Somes RG 1970 The influence of the rate of feathering allele
Kn on various quantitative traits in chickens. Poult Sci 49(5):1251-1256.
Warren DC 1976 Feather-sexing chicks. Poult Tribune 82(2):32-34.
Weissmann A, Reitemeier S, Hahn A, Gottschalk J, Einspanier A 2013 Sexing domestic chicken before hatch: a new method for
in ovo gender identification. Theriogenology 80(3):199-205.